

by Jake Dalzell, Hazel Garrett, Catriona Forrest, Wayne Liang, Rosalind Mackey, Denis Pavlov, and Josh Simpson
This was a small project we threw together over three days on our Plant Sciences fieldtrip to Portugal. Everyone on the fieldtrip used six different techniques to explore plant physiology and ecology, and each group came up with a research question that could be answered using some of these techniques. Given the short time frame, our samples were biased and small (too small for robust statistics!). We can’t really draw any hard conclusions from our results, but they do show interesting trends.
Our project compared the strategies used by two parasitic plants in the Orobanchaceae and a shared host species in the Fabaceae. We also compared host individuals which were parasitised by each species with each other and with uninfected hosts. We were particularly interested in how water use differed between the different plants. We expected to see a lower water potential in the parasites than in their host, as has been shown in other Orobanchaceae︎¹,². This water potential gradient generates a sink, so there is a flux of xylem fluid from host roots to parasite¹.
Study species
We chose to study a holoparasite, Phelipanche ramosa; a hemiparasite, Bellardia trixago; and a legume that they both parasitise, Medicago polymorpha. These were abundant in the grassland where we were staying, Quinta de São Pedro.




Mycorrhizal staining
We cleared the roots using Sodium hydroxide, then stained for mycorrhizal fungi by using a blue dye that binds to chitin, followed by washing with Hydrochloric acid and then water.


The results were fairly clear: the parasites had minimal fungal infection (holoparasite) or none (hemiparasite), while the host had many hyphae and arbuscules. The hyphae visible in the holoparasite may have been an early stage of mycorrhization but the way the samples were prepared means the context of the root was lost. Fungal structures are visible in blue:


| Host | Holoparasite | Hemiparasite | |
| Hyphae? | Yes | Yes | No |
| Arbuscules? | Yes | No | No |
The lack of mycorrhizae in the parasites suggests that they do not need fungi to acquire resources as they can just steal them (and hence the carbon cost of mycorrhizae is not worth it). The resources are acquired from mycorrhizae at a carbon cost by the host, and then promptly stolen by the parasites. It would be interesting to compare with a hemiparasite grown without a host – does it now form mycorrhizae, as it cannot steal resources from other plants?
Water relations
Licor was used to measure stomatal conductance and other metrics, and estimate water use efficiency. A pressure bomb was used to measure instantaneous water potential in stems cut around 3cm from the base. The holoparasite could not be measured using licor as it does not have substantial leaves.
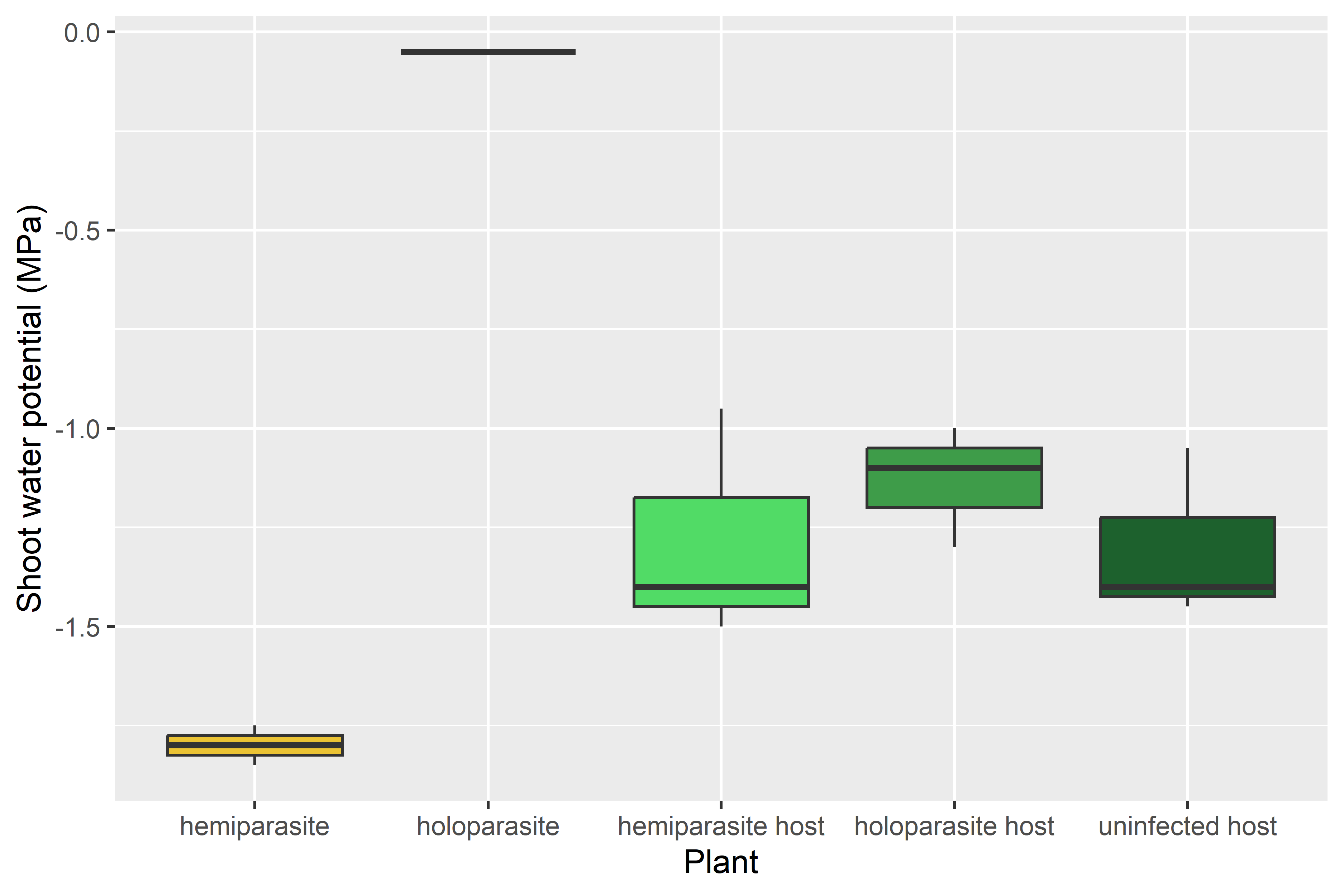
The shoot water potential was much lower in the hemiparasite than in the host, as expected. The holoparasite had a very high water potential, which was unexpected. This may have been due to issues in measurement, as the texture of the stems made gauging the point at which water was visible at the cut end difficult. It may be that the water potential in the tubercle of the holoparasite was lower as in Cistanche², so it could still act as a sink on the host plant. The extremely high water potential is still strange though, and merits further research.
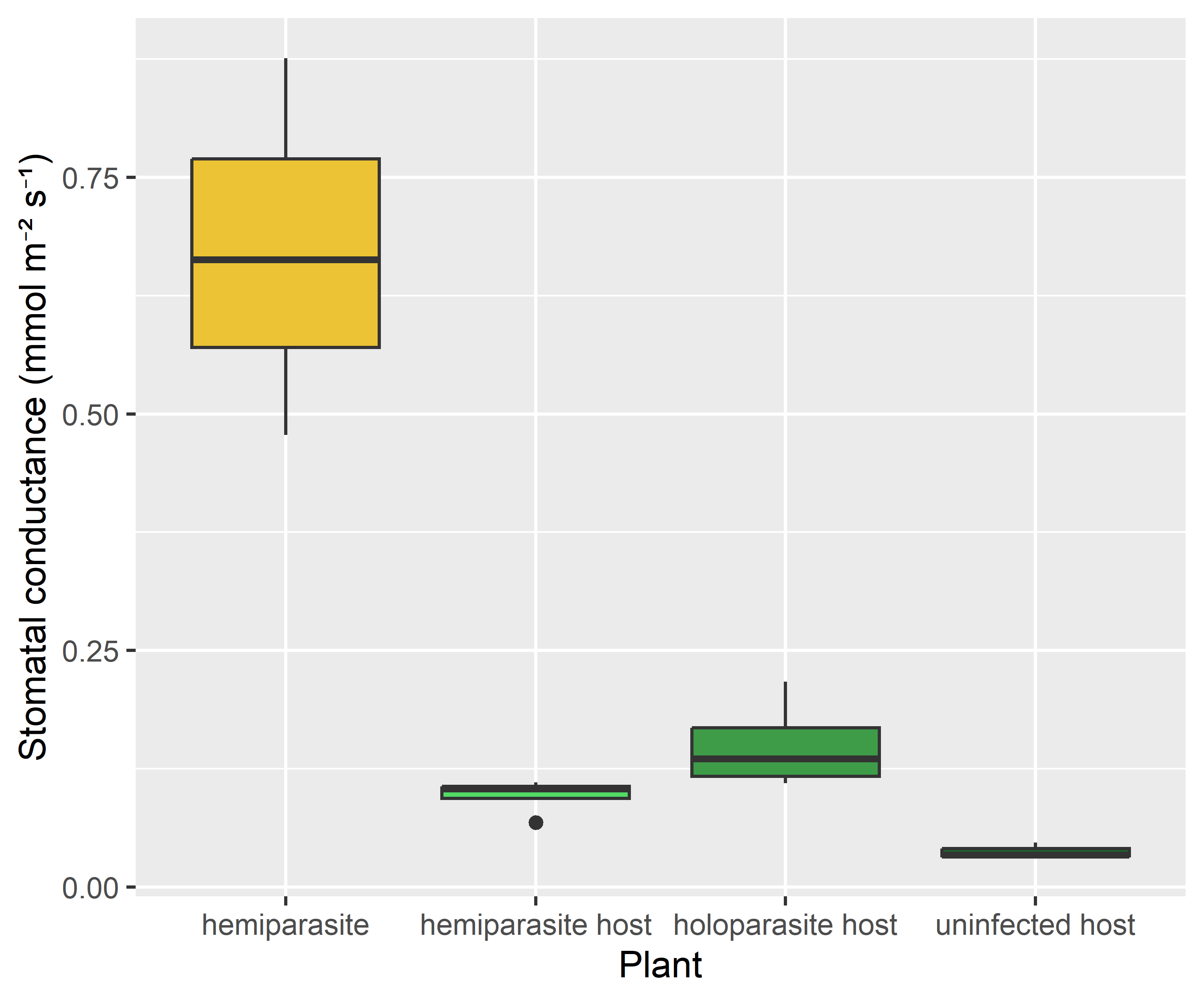
Stomatal conductance was considerably higher in the hemiparasite than in the host, providing a mechanism for its low water potential.
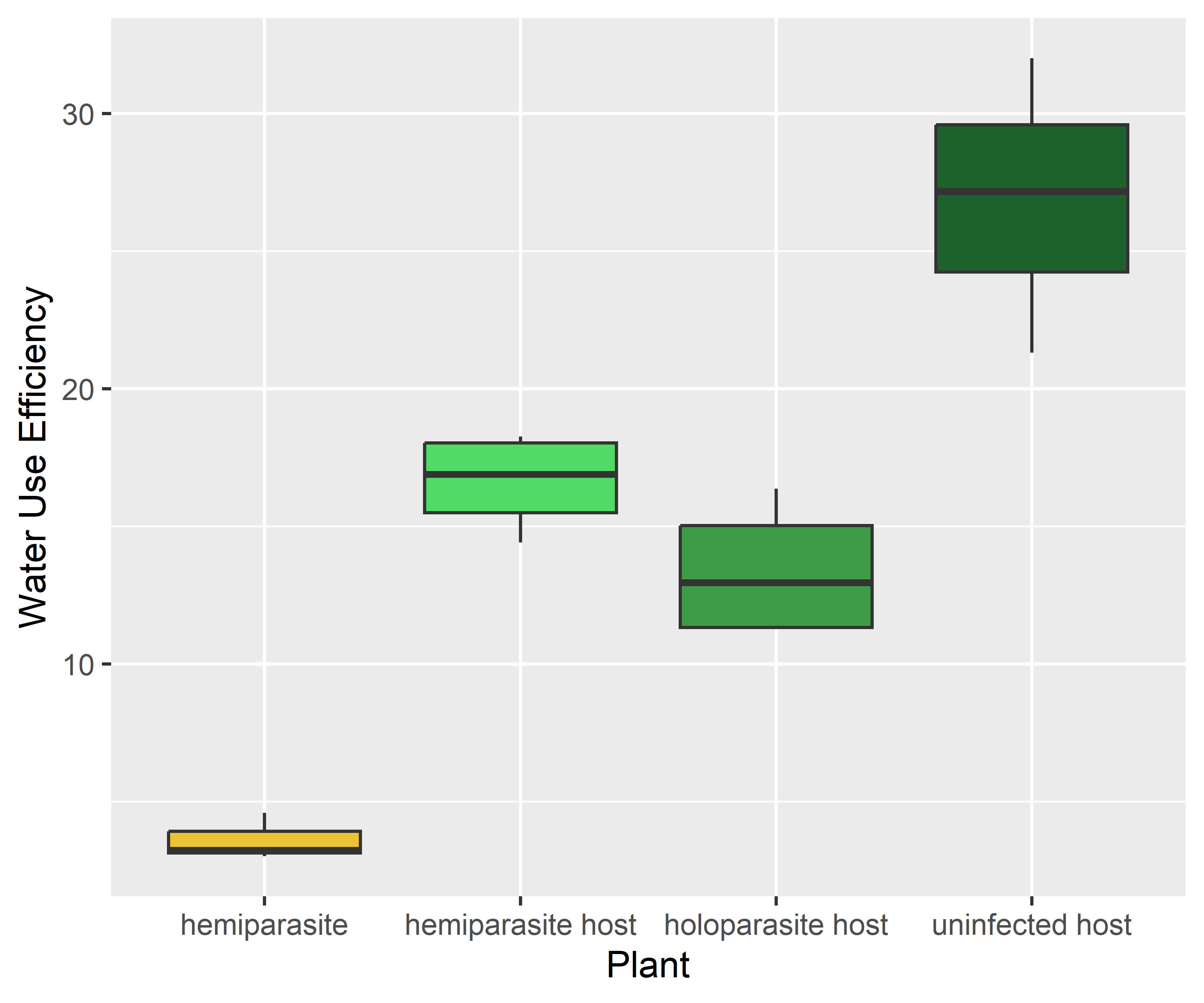
The water use efficiency of the hemiparasite was much lower than that of the host. This can be explained by its high conductance.
Overall the hemiparasite has a strong “spender” strategy, freely using resources that it gains from its host and transpiring a lot of water to pull a low water potential and get more host resources.
References
- Ackroyd, Robert D., and Jonathan D. Graves. 1997. ‘The Regulation of the Water Potential Gradient in the Host and Parasite Relationship between Sorghum bicolor and Striga hermonthica’. Annals of Botany 80 (5): 649–56. https://doi.org/10.1006/anbo.1997.0506.
- Fahmy, Gamal Mohammad. 2013. ‘Ecophysiology of the Holoparasitic Angiosperm Cistanche phelypaea (Orobanchaceae) in a Coastal Salt Marsh’. Turkish Journal of Botany 37: 908–19. https://doi.org/10.3906/bot-1210-48.
Leave a Reply